")
 Scheme")
L-Arabinose/D-Galactose Assay Kit
115 assays (manual) / 1150 assays (microplate) / 1150 assays (auto-analyser)
Long term stability: See individual component labels
0.69 mg/L (D-galactose)
~ 6 min (D-galactose)
Advantages
- Extended cofactors stability. Dissolved cofactors stable for > 1 year at 4oC.
- Very rapid reaction due to inclusion of galactose mutarotase (patented technology)
- Very cost effective
- All reagents stable for > 2 years after preparation
- Only enzymatic kit available
- Mega-Calc™ software tool is available from our website for hassle-free raw data processing
- Standard included
- Suitable for manual, microplate and auto-analyser formats
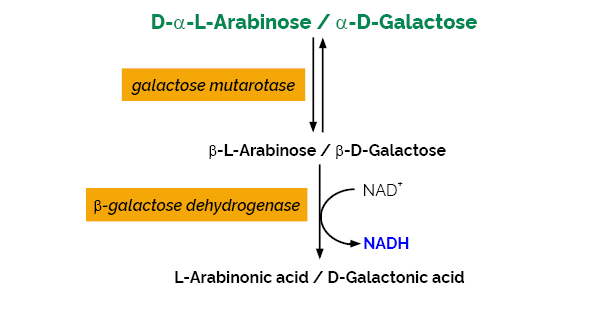
The L-Arabinose/D-Galactose test kit is a simple, reliable and accurate UV method for the measurement and analysis of L-arabinose and/or D-galactose in various materials including foods, feeds, beverages and plant products.
Note for Content: The number of manual tests per kit can be doubled if all volumes are halved. This can be readily accommodated using the MegaQuantTM Wave Spectrophotometer (D-MQWAVE).
See more of our monosaccharide assay kits.
Related Products
Similar Products
Related Carbohydrates
Associated Enzymes
Publications

Measurement of available carbohydrates in cereal and cereal products, dairy products, vegetables, fruit and related food products and animal feeds: First Action 2020.07.
McCleary, B. V. & McLoughlin, C. (2021). Journal of AOAC International, qsab019.
Measurement of available carbohydrates in cereal and cereal products, dairy products, vegetables, fruit and related food products and animal feeds: First Action 2020.07.
McCleary, B. V. & McLoughlin, C. (2021). Journal of AOAC International, qsab019.
Background: The level of available carbohydrates in our diet is directly linked to two major diseases; obesity and Type II diabetes. Despite this, to date there is no method available to allow direct and accurate measurement of available carbohydrates in human and animal foods. Objective: The aim of this research was to develop a method that would allow simple and accurate measurement of available carbohydrates, defined as non-resistant starch, maltodextrins, maltose, isomaltose, sucrose, lactose, glucose, fructose and galactose. Method: Non-resistant (digestible) starch is hydrolysed to glucose and maltose by pancreatic α-amylase and amyloglucosidase at pH 6.0 with shaking or stirring at 37°C for 4 h. Sucrose, lactose, maltose and isomaltose are completely hydrolyzed by specific enzymes to their constituent monosaccharides, which are then measured using pure enzymes in a single reaction cuvette. Results: A method has been developed that allows the accurate measurement of available carbohydrates in all cereal, vegetable, fruit, food, and feed products, including dairy products. Conclusions: A single-laboratory validation was performed on a wide range of food and feed products. The inter-day repeatability (%RSDr) was <3.58% (w/w) across a range of samples containing 44.1 to 88.9% available carbohydrates. The LOD and LOQ obtained were 0.054% (w/w) and 0.179% (w/w), respectively. The method is all inclusive, specific, robust and simple to use. Highlights: A unique method has been developed for the direct measurement of available carbohydrates, entailing separate measurement of glucose, fructose and galactose; information of value in determining the glycemic index of foods.
McCleary, B. V., McKie, V. A., Draga, A., Rooney, E., Mangan, D. & Larkin, J. (2015). Carbohydrate Research, 407, 79-96.
McCleary, B. V., McKie, V. A., Draga, A., Rooney, E., Mangan, D. & Larkin, J. (2015). Carbohydrate Research, 407, 79-96.
A range of α-L-arabinofuranosyl-(1-4)-β-D-xylo-oligosaccharides (AXOS) were produced by hydrolysis of wheat flour arabinoxylan (WAX) and acid debranched arabinoxylan (ADWAX), in the presence and absence of an AXH-d3 α-L-arabinofuranosidase, by several GH10 and GH11 β-xylanases. The structures of the oligosaccharides were characterised by GC-MS and NMR and by hydrolysis by a range of α-L-arabinofuranosidases and β-xylosidase. The AXOS were purified and used to characterise the action patterns of the specific α-L-arabinofuranosidases. These enzymes, in combination with either Cellvibrio mixtus or Neocallimastix patriciarum β -xylanase, were used to produce elevated levels of specific AXOS on hydrolysis of WAX, such as 32-α-L-Araf-(1-4)-β-D-xylobiose (A3X), 23-α-L-Araf-(1-4)-β-D-xylotriose (A2XX), 33-α-L-Araf-(1-4)-β-D-xylotriose (A3XX), 22-α-L-Araf-(1-4)-β-D-xylotriose (XA2X), 32-α-L-Araf (1-4)-β-D-xylotriose (XA3X), 23-α-L-Araf-(1-4)-β-D-xylotetraose (XA2XX), 33-α-L-Araf-(1-4)-β-D-xylotetraose (XA3XX), 23 ,33-di-α-L-Araf-(1-4)-β-D-xylotriose (A2+3XX), 23,33-di-α-L-Araf-(1-4)-β-D-xylotetraose (XA2+3XX), 24,34-di-α-L-Araf-(1-4)-β-D-xylopentaose (XA2+3XXX) and 33,34-di-α-L-Araf-(1-4)-β-D-xylopentaose (XA3A3XX), many of which have not previously been produced in sufficient quantities to allow their use as substrates in further enzymic studies. For A2,3XX, yields of approximately 16% of the starting material (wheat arabinoxylan) have been achieved. Mixtures of the α-L-arabinofuranosidases, with specific action on AXOS, have been combined with β-xylosidase and β-xylanase to obtain an optimal mixture for hydrolysis of arabinoxylan to L-arabinose and D-xylose.
Understanding the substrate recognition and catalytic mechanism of methyl fucosidases from glycoside hydrolase family 139.
McIver, Z., Moraleda-Montoya, A., Chen, Z., Epa, R., Starns, D., Davy, M., Garcia-Alija, M., Basle, A., Schubert, M., Ndeh, D., Trastoy, B., Williama, S. J., Guerin, M. E. & Cartmell, A. Journal of Biological Chemistry, 110407.
Understanding the substrate recognition and catalytic mechanism of methyl fucosidases from glycoside hydrolase family 139.
McIver, Z., Moraleda-Montoya, A., Chen, Z., Epa, R., Starns, D., Davy, M., Garcia-Alija, M., Basle, A., Schubert, M., Ndeh, D., Trastoy, B., Williama, S. J., Guerin, M. E. & Cartmell, A. Journal of Biological Chemistry, 110407.
Rhamnogalacturonan II is one of the most complex plant cell wall carbohydrates and is composed of 13 different sugars and 21 different glycosidic linkages. It is abundant in fruit and indulgence foods, such as chocolate and wine, making it common in the human diet. The human colonic commensal Bacteroides thetaiotaomicron expresses a consortium of 22 enzymes to metabolise rhamnogalacturonan II, some of which exclusively target sugars unique to rhamnogalacturonan II. Several of these enzyme families remain poorly described, and, consequently, our knowledge of rhamnogalacturonan II metabolism is limited. Chief among the poorly understood activities is glycoside hydrolase (GH) family 139, with targets α1,2-2O-methyl L-fucoside linkages, a sugar residue a sugar not found in any other plant cell wall complex glycans. Although the founding enzyme BT0984 was placed in the RG-II degradative pathway, no GH139 structure or catalytic blueprint had been available. We report the crystal structures of BT0984 and a second homologue, and reveal that the family operates with inverting stereochemistry. Using this data we undertook a mutagenic strategy, backed by molecular dynamics, to identify the important substrate binding and catalytic residues, mapping these residues throughout the GH139 family revealing the importance of the O2 methyl interaction of the substrate. We propose a catalytic mechanism that uses a non-canonical Asn as a catalytic base and shares similarity with L-fucosidases/L-galactosidases of family GH95.
A time-course analysis of Aspergillus terreus secretomes reveals the importance of pectin-degrading enzymes to increase the digestibility of soybean meal.
Plouhinec, L., Bonnin, E., Kielbasa, M., Armengaud, J., Neugnot, V., Berrin, J. G. & Lafond, M. (2024). Applied and Environmental Microbiology, 90(9), e02153-23.
A time-course analysis of Aspergillus terreus secretomes reveals the importance of pectin-degrading enzymes to increase the digestibility of soybean meal.
Plouhinec, L., Bonnin, E., Kielbasa, M., Armengaud, J., Neugnot, V., Berrin, J. G. & Lafond, M. (2024). Applied and Environmental Microbiology, 90(9), e02153-23.
Considering an ever-growing global population, which hit 8 billion people in the fall of 2022, it is essential to find solutions to avoid croplands competition between human food and animal feed. Agricultural co-products such as soybean meals have become important components of the circular economy thanks to their use in animal feed. Their implementation was made possible by the addition of exogenous enzymes in the diet of monogastric animals, especially fungal carbohydrate-active enzymes (CAZymes). Here, we describe a time-course production and analysis of Aspergillus terreus secretomes for the identification of CAZymes able to enhance the digestibility of soybean meals. Functional assays revealed that the release of nutrients and the degradation of pectins in soybean meals can be tightly interconnected. Using a comparative proteomics approach, we identified several fungal pectin-degrading enzymes leading to increased assimilable nutrients in the soluble fraction of soybean meals. Our results reinforce the importance of deconstructing pectic polysaccharides in feedstuffs and contribute to sharpen our understanding of the fungal enzymatic interplays involved in pectin hydrolysis.IMPORTANCEIn the present study, we developed a strategy to identify the key fungal enzymatic activities involved in the improvement of soybean meal (SBM) digestibility. Our data unravel the importance of pectin degradation for the release of nutrients from SBM and provide some insights regarding the degradation of rhamnogalacturonan-I (RG-I) by ascomycetes. Indeed, the hydrolysis of pectins and RG-I by human microbiota is well documented in the literature, but our knowledge of the fungal CAZymes at play for the degradation of soybean pectins remains hitherto underexplored. Due to its wide use in animal feed, improving the digestibility of SBM by enzymatic treatments is a current challenge for feed additive suppliers. Since non-starch polysaccharides and pectins have often been reported for their anti-nutritional role in SBM, we believe this study will provide new avenues toward the improvement of enzymatic cocktails for animal nutrition and health.
Chemical composition and bioactivity of oilseed cake extracts obtained by subcritical and modified subcritical water.
Švarc-Gajić, J., Rodrigues, F., Moreira, M. M., Delerue-Matos, C., Morais, S., Dorosh, O., Silva, A. M., Bassani, A., Dzedik, V. & Spigno, G. (2022). Bioresources and Bioprocessing, 9(1), 1-14.
Chemical composition and bioactivity of oilseed cake extracts obtained by subcritical and modified subcritical water.
Švarc-Gajić, J., Rodrigues, F., Moreira, M. M., Delerue-Matos, C., Morais, S., Dorosh, O., Silva, A. M., Bassani, A., Dzedik, V. & Spigno, G. (2022). Bioresources and Bioprocessing, 9(1), 1-14.
Recovery of bioactive compounds from biowaste is gaining more and more interest in circular economy models. The oilseed cakes are usually insufficiently exploited by most technologies since they represent valuable matrices abundant in proteins, minerals, and phytochemicals, but their use is mostly limited to feed ingredients, fertilizers or biofuel production. This study was thus focused on the exploration of new valorization pathways of oilseed cakes by subcritical water, representing a safe and economic alternative in the creation of value chains. Pumpkin, hemp, and flax seed cakes were treated with subcritical water in nitrogen and carbon-dioxide atmospheres, as well as in nitrogen atmosphere with the addition of acid catalyst. The degradation of carbohydrate fraction was studied by quantifying sugars and sugar degradation products in the obtained extracts. The extracts obtained under different conditions were further compared chemically with respect to total phenols and flavonoids, as well as to the content of individual phenolic compounds. Furthermore, the effects of subcritical water treatment conditions on antioxidant, antiradical and cytotoxic properties of thus obtained extracts were defined and discussed.
High-pressure autohydrolysis process of wheat straw for cellulose recovery and subsequent use in PBAT composites preparation.
Fiorentini, C., Bassani, A., Garrido, G. D., Merino, D., Perotto, G., Athanassiou, A., Prantie, J., Halonen, N. & Spigno, G. (2022). Biocatalysis and Agricultural Biotechnology, 39, 102282.
High-pressure autohydrolysis process of wheat straw for cellulose recovery and subsequent use in PBAT composites preparation.
Fiorentini, C., Bassani, A., Garrido, G. D., Merino, D., Perotto, G., Athanassiou, A., Prantie, J., Halonen, N. & Spigno, G. (2022). Biocatalysis and Agricultural Biotechnology, 39, 102282.
The effect of autohydrolysis (AH) temperature (165°C, 195°C, 225°C) on the structure, purity, and recovery yield of the cellulose residue isolated after additional alkaline and bleaching steps from wheat straw, was investigated. The processes were quantified for mass yields in the different steps and for antioxidants and sugars release during AH. AH at 195°C allowed for the highest cellulose residue yield (83.5%) with purity (~70%) and structure similar to the other residues. FTIR and XRD analyses showed straw cellulose (SC) with a type II polymorphism and crystallinity index increasing with AH temperature. SC obtained at the end of the entire fractionation process (SC-195°C) starting from AH residue-195°C was tested as a reinforcing agent in different percentage (0, 2 and 5% by weight) in poly(butylene adipate-co-terephthalate) (PBAT) films. The Young's modulus of the films increased by ~17% with 5 wt% cellulose, while tensile strength and elongation at break decreased.
Utilization of stalk waste separated during processing of sun-dried figs (Ficus carica) as a source of pectin: Extraction and determination of molecular and functional properties.
Çavdaroğlu, E. & Yemenicioğlu, A. (2021). LWT, 154, 112624.
Utilization of stalk waste separated during processing of sun-dried figs (Ficus carica) as a source of pectin: Extraction and determination of molecular and functional properties.
Çavdaroğlu, E. & Yemenicioğlu, A. (2021). LWT, 154, 112624.
This study aimed the utilization of fig stalk waste as an alternative pectin source. For this purpose, the characteristics of extracted stalk waste pectin (SP) were compared with those of citrus pectin (CP) and pectin extracted from defected substandard whole sun-dried figs (FP). The SP had a higher extraction yield (11.7%) than FP (9.4%). The galacturonic acid content and degree of esterification of SP (32.3 and 50.1%) were higher than those of FP (19.9 and 38.8%), but lower than those of CP (79.3 and 56.2%), respectively. The SP and CP had different sugar compositions (d-glucose, l-rhamnose, d-galactose and l-arabinose) and weight average molecular weights, but similar FTIR profiles. The SP showed almost 1.9 and 1.6-fold higher Trolox equivalent antioxidant capacity (TEAC), and 2.7 and 2.0-fold higher water absorption capacity than CP and FP, respectively. SP at 3% (w/w) showed the second highest viscosity after CP and the highest emulsion stability. Gels of SP and CP at 1.75–3% range had similar firmness, but SP formed more fracturable gels than CP. Sun-dried fig stalk waste is a better source of pectin than defected substandard whole sun-dried figs. The SP could be utilized to develop functional food with alternative textural and rheological properties.
Escherichia coli AraJ boosts utilization of arabinose in metabolically engineered cyanobacterium Synechocystis sp. PCC 6803.
Ranade, S. & He, Q. (2021). AMB Express, 11(1), 1-14.
Escherichia coli AraJ boosts utilization of arabinose in metabolically engineered cyanobacterium Synechocystis sp. PCC 6803.
Ranade, S. & He, Q. (2021). AMB Express, 11(1), 1-14.
Lignocellulosic biomass can serve as an inexpensive and renewable source of carbon for the biosynthesis of commercially important compounds. L-arabinose is the second most abundant pentose sugar present in the plant materials. Model cyanobacterium Synechocystis sp. PCC 6803 is incapable of catabolism of L-arabinose as a source of carbon and energy. In this study, all the heterologous genes expressed in Synechocystis were derived from Escherichia coli K-12. Initially we constructed four Synechocystis strains that expressed AraBAD enzymes involved in L-arabinose catabolism, either in combination with or without one of the three arabinose transporters, AraE, AraFGH or AraJ. Among the recombinants, the strain possessing AraJ transporter was observed to be the most efficient in terms of dry biomass production and L-arabinose consumption. Later, an additional strain was generated by the expression of AraJ in the AraE-possessing strain. The resultant strain was shown to be advantageous over its parent. This study demonstrates that AraJ, a protein with hitherto unknown function plays a role in the uptake of L-arabinose to boost its catabolism in the transgenic Synechocystis strains. The work also contributes to the current knowledge regarding metabolic engineering of cyanobacteria for the utilization of pentose sugars.
The protective effect of Scenedesmus dimorphus polysaccharide as an antioxidant and antiaging agent on aging rat model induced by D-galactose.
Armaini, A. & Imelda, I. (2021). Journal of Applied Pharmaceutical Science, 11(05), 054-063.
The protective effect of Scenedesmus dimorphus polysaccharide as an antioxidant and antiaging agent on aging rat model induced by D-galactose.
Armaini, A. & Imelda, I. (2021). Journal of Applied Pharmaceutical Science, 11(05), 054-063.
Premature skin aging occurs due to the increased formation of reactive oxygen species (ROS), which causes oxidative stress, DNA damage, and collagen degradation. This study investigates the protective effect of Scenedesmus dimorphus polysaccharides (SDP) as an antioxidant and anti-aging agent on an aging rat model induced by D-galactose (D-gal). This study used 48 male Wistar rats divided into six groups: (1) normal control, (2) polysaccharide control, (3) aging control induced by D-gal 0.25 mg/g bw/days, (4) drug control (vitamin E) treated twice with SDP in an aging rat model, (5) D-gal + SDP (0.2 mg/g bw), and (6) D-gal + SDP (0.8 mg/g bw) treated with oral treatment and observed for periods of 2, 4, and 8 weeks. Giving SDP at a dose of 0.8 mg/g bw can increase superoxide dismutases and catalase activity and reduce malondialdehyde after 8 weeks of observation, which is better than giving vitamin E. The treatment of SDP can stimulate collagen synthesis and reduce advanced glycation end products. Histopathology shows an increase in the area of fibrocollagen proportions and deposition from the matrix on giving SDP, which is better than vitamin E, since SDP can repair skin tissue. Thus, SDP can be useful as an antioxidant and anti-aging agent in an aging Wistar rats model.
Monosaccharide constituents of potato root exudate influence hatching of the white potato cyst nematode.
Bell, C. A., Mobayed, W., Lilley, C. J. & Urwin, P. (2021). PhytoFrontiers, 1-26.
Monosaccharide constituents of potato root exudate influence hatching of the white potato cyst nematode.
Bell, C. A., Mobayed, W., Lilley, C. J. & Urwin, P. (2021). PhytoFrontiers, 1-26.
Plants secrete a large array of compounds into the rhizosphere to facilitate interactions with their biotic environment. Some of these exuded-compounds stimulate the hatching of obligate plant-parasitic nematodes, ultimately leading to a detrimental effect on the host plant. Determining these cues can help to provide new mechanisms for control and aid nematode management schemes. Here we show that glucose, fructose and arabinose, which are all present in potato root exudate (PRE), induce hatching of white potato cyst nematode (Globodera pallida) eggs whereas five other PRE-sugars had no effect. Although these monosaccharides resulted in significant hatching none induced the same level as PRE, suggesting that other components, possibly in combination, contribute to stimulation of nematode hatching. Glucose, but not arabinose or fructose, was also observed to attract juvenile G. pallida, indicating that these hatch-inducing components can have different roles in different stages of the life cycle. Applying a solution of these monosaccharides to G. pallida-infested soil pre-potato planting initiated hatching in the absence of a host. Host absence resulted in nematode mortality and a reduction in the G. pallida population. Therefore, subsequent invasion of the crop post-planting was also reduced, compared to untreated soil. Our data suggest that monosaccharide components of PRE play an important role in the hatching and attraction of G. pallida. As a result the hatch-inducing monosaccharides can be applied as a pre-planting treatment to induce hatching and reduce subsequent infection rates.
A new, quick, and simple protocol to evaluate microalgae polysaccharide composition.
Decamp, A., Michelo, O., Rabbat, C., Laroche, C., Grizeau, D., Pruvost, J. & Gonçalves, O. (2021). Marine Drugs, 19(2), 101.
A new, quick, and simple protocol to evaluate microalgae polysaccharide composition.
Decamp, A., Michelo, O., Rabbat, C., Laroche, C., Grizeau, D., Pruvost, J. & Gonçalves, O. (2021). Marine Drugs, 19(2), 101.
In this work, a new methodological approach, relying on the high specificity of enzymes in a complex mixture, was developed to estimate the composition of bioactive polysaccharides produced by microalgae, directly in algal cultures. The objective was to set up a protocol to target oligomers commonly known to be associated with exopolysaccharides’ (EPS) nutraceutical and pharmaceutical activities (i.e., rhamnose, fucose, acidic sugars, etc.) without the constraints classically associated with chromatographic methods, while maintaining a resolution sufficiently high to enable their monitoring in the culture system. Determination of the monosaccharide content required the application of acid hydrolysis (2 M trifluoroacetic acid) followed by NaOH (2 M) neutralization. Quantification was then carried out directly on the fresh hydrolysate using enzyme kits corresponding to the main monosaccharides in a pre-determined composition of the polysaccharides under analysis. Initial results showed that the enzymes were not sensitive to the presence of TFA and NaOH, so the methodology could be carried out on fresh hydrolysate. The limits of quantification of the method were estimated as being in the order of the log of nanograms of monosaccharides per well, thus positioning it among the chromatographic methods in terms of analytical performance. A comparative analysis of the results obtained by the enzymatic method with a reference method (high-performance anion-exchange chromatography) confirmed good recovery rates, thus validating the closeness of the protocol. Finally, analyses of raw culture media were carried out and compared to the results obtained in miliQ water; no differences were observed. The new approach is a quick, functional analysis method allowing routine monitoring of the quality of bioactive polysaccharides in algal cultures grown in photobioreactors.
A novel glycosidase plate-based assay for the quantification of galactosylation and sialylation on human IgG.
Rebello, O. D., Gardner, R. A., Urbanowicz, P. A., Bolam, D. N., Crouch, L. I., Falck, D. & Spencer, D. I. (2020). Glycoconjugate Journal, 37(6), 691-702.
A novel glycosidase plate-based assay for the quantification of galactosylation and sialylation on human IgG.
Rebello, O. D., Gardner, R. A., Urbanowicz, P. A., Bolam, D. N., Crouch, L. I., Falck, D. & Spencer, D. I. (2020). Glycoconjugate Journal, 37(6), 691-702.
Changes in human IgG galactosylation and sialylation have been associated with several inflammatory diseases which are a major burden on the health care system. A large body of work on well-established glycomic and glycopeptidomic assays has repeatedly demonstrated inflammation-induced changes in IgG glycosylation. However, these assays are usually based on specialized analytical instrumentation which could be considered a technical barrier for uptake by some laboratories. Hence there is a growing demand for simple biochemical assays for analyzing these glycosylation changes. We have addressed this need by introducing a novel glycosidase plate-based assay for the absolute quantification of galactosylation and sialylation on IgG. IgG glycoproteins are treated with specific exoglycosidases to release the galactose and/or sialic acid residues. The released galactose monosaccharides are subsequently used in an enzymatic redox reaction that produces a fluorescence signal that is quantitative for the amount of galactosylation and, in-turn, sialylation on IgG. The glycosidase plate-based assay has the potential to be a simple, initial screening assay or an alternative assay to the usage of high-end analytical platforms such as HILIC-FLD-MSn when considering the analysis of galactosylation and sialylation on IgG. We have demonstrated this by comparing our assay to an industrial established HILIC-FLD-MSn glycomic analysis of 15 patient samples and obtained a Pearson’s r correlation coefficient of 0.8208 between the two methods.
Structural analysis of β‐L‐arabinobiose‐binding protein in the metabolic pathway of hydroxyproline‐rich glycoproteins in Bifidobacterium longum.
Miyake, M., Terada, T., Shimokawa, M., Sugimoto, N., Arakawa, T., Shimizu, K., Igarashi, K., Fujita, K. & Fushinobu, S. (2020). The FEBS Journal, 287(23), 5114-5129.
Structural analysis of β‐L‐arabinobiose‐binding protein in the metabolic pathway of hydroxyproline‐rich glycoproteins in Bifidobacterium longum.
Miyake, M., Terada, T., Shimokawa, M., Sugimoto, N., Arakawa, T., Shimizu, K., Igarashi, K., Fujita, K. & Fushinobu, S. (2020). The FEBS Journal, 287(23), 5114-5129.
Bifidobacterium longum is a symbiotic human gut bacterium that has a degradation system for β‐arabinooligosaccharides, which are present in the hydroxyproline‐rich glycoproteins of edible plants. Whereas microbial degradation systems for α‐linked arabinofuranosyl carbohydrates have been extensively studied, little is understood about the degradation systems targeting β‐linked arabinofuranosyl carbohydrates. We functionally and structurally analyzed a substrate‐binding protein (SBP) of a putative ABC transporter (BLLJ_0208) in the β‐arabinooligosaccharide degradation system. Thermal shift assays and isothermal titration calorimetry revealed that the SBP specifically bound Araf‐β1,2‐Araf (β‐Ara2) with a Kd of 0.150 μm, but did not bind L‐arabinose or methyl‐β‐Ara2. Therefore, the SBP was termed β‐arabinobiose‐binding protein (BABP). Crystal structures of BABP complexed with β‐Ara2 were determined at resolutions of up to 1.78 Å. The findings showed that β‐Ara2 was bound to BABP within a short tunnel between two lobes as an α‐anomeric form at its reducing end. BABP forms extensive interactions with β‐Ara2, and its binding mode was unique among SBPs. A molecular dynamics simulation revealed that the closed conformation of substrate‐bound BABP is stable, whereas substrate‐free form can adopt a fully open and two distinct semi‐open states. The importer system specific for β‐Ara2 may contribute to microbial survival in biological niches with limited amounts of digestible carbohydrates.
Comparison of Japanese and Indian intestinal microbiota shows diet-dependent interaction between bacteria and fungi.
Pareek, S., Kurakawa, T., Das, B., Motooka, D., Nakaya, S., Rongsen-Chandola, T. et al. (2019). NPJ Biofilms and Microbiomes, 5(1), 1-13.
Comparison of Japanese and Indian intestinal microbiota shows diet-dependent interaction between bacteria and fungi.
Pareek, S., Kurakawa, T., Das, B., Motooka, D., Nakaya, S., Rongsen-Chandola, T. et al. (2019). NPJ Biofilms and Microbiomes, 5(1), 1-13.
The bacterial species living in the gut mediate many aspects of biological processes such as nutrition and activation of adaptive immunity. In addition, commensal fungi residing in the intestine also influence host health. Although the interaction of bacterium and fungus has been shown, its precise mechanism during colonization of the human intestine remains largely unknown. Here, we show interaction between bacterial and fungal species for utilization of dietary components driving their efficient growth in the intestine. Next generation sequencing of fecal samples from Japanese and Indian adults revealed differential patterns of bacterial and fungal composition. In particular, Indians, who consume more plant polysaccharides than Japanese, harbored increased numbers of Prevotella and Candida. Candida spp. showed strong growth responses to the plant polysaccharide arabinoxylan in vitro. Furthermore, the culture supernatants of Candida spp. grown with arabinoxylan promoted rapid proliferation of Prevotella copri. Arabinose was identified as a potential growth-inducing factor in the Candida culture supernatants. Candida spp. exhibited a growth response to xylose, but not to arabinose, whereas P. copri proliferated in response to both xylose and arabinose. Candida spp., but not P. copri, colonized the intestine of germ-free mice. However, P. copri successfully colonized mouse intestine already harboring Candida. These findings demonstrate a proof of concept that fungal members of gut microbiota can facilitate a colonization of the intestine by their bacterial counterparts, potentially mediated by a dietary metabolite.
Cross-linking of diluted alkali-soluble pectin from apple (Malus domestica fruit) in different acid-base conditions.
Gawkowska, D., Cieśla, J., Zdunek, A. & Cybulska, J. (2019). Food Hydrocolloids, 92, 285-292.
Cross-linking of diluted alkali-soluble pectin from apple (Malus domestica fruit) in different acid-base conditions.
Gawkowska, D., Cieśla, J., Zdunek, A. & Cybulska, J. (2019). Food Hydrocolloids, 92, 285-292.
A diluted alkali-soluble pectin (DASP) fraction, extracted using sodium carbonate, is characterized by a low degree of methylesterification and has the ability to self-organize on mica. The aim of this study was to characterize the cross-linking process of this fraction, extracted from apples, over a wide pH range (3-11) and without the addition of salt. An FT-IR study showed an increase in the intensity of bands connected with νas and νs (COO−) and a decrease in the intensity of the band associated with ν (C=O) in the carboxyl group with increasing pH, which indicated the dissociation of the carboxyl groups of galacturonic acid units. An increase in the surface electrical charge of particles in the pH range of 3-7 confirmed this. The value of the average apparent dissociation constant (∼4.60) indicated the acidic character of the DASP fraction. An AFM study showed the morphological changes of the DASP fraction with increasing pH, which allowed for the evaluation of the cross-linking process. This fraction formed a network on mica at pH 4 and 9, while the aggregates were noted mainly at pH 11. For totally ionized carboxyl groups (pH 7), the pectin chains were separated from each other due to the electrostatic repulsion between them.
Safety Information
Download Safety Data Sheet
Select a specific region and the Safety Data Sheet will open in a new window. Please make sure your browser doesn’t block pop-up windows

")
 (R-BAMR3)")
 (E-ANAAM)")
")
")
 (E-GALDH)")



