")
 Scheme")
myo-Inositol Assay Kit
50 assays per kit
Long term stability: See individual component labels
Advantages
- Very cost effective
- All reagents stable for > 2 years after preparation
- Only enzymatic kit available
- Rapid reaction
- Mega-Calc™ software tool is available from our website for hassle-free raw data processing
- Standard included
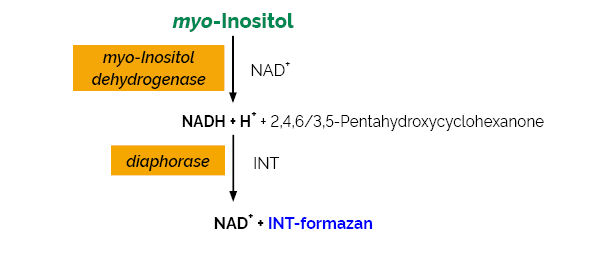
The myo-Inositol Assay Kit is a reliable and accurate enzymatic UV-method for the specific measurement and analysis of myo-inositol in animal feeds, foods and various other materials.
Phytic acid content of samples with very low levels of free myo-inositol can also be determined using K-INOSL. This can be achieved by measuring the amount of myo-inositol released after de-phosphorylation of phytic acid with the enzymes phytase and alkaline phosphatase, as used with the Megazyme Phytic Acid/Total Phosphorus Assay Kit (K-PHYT).
Not suitable for the determination of myo-inositol in baby formula.
Note for Content: The number of manual tests per kit can be doubled if all volumes are halved. This can be readily accommodated using the MegaQuantTM Wave Spectrophotometer (D-MQWAVE).
Find out more of our alcohol test kits.
Related Products
Publications
SlMIPS2, a myo-inositol phosphate synthase, regulates phosphate homeostasis by influencing SlPHL1 and SlSPX2 levels in tomato seedlings.
Roychowdhury, A., Kaushik, D., Bhosale, J. B., Srivastava, R. & Kumar, R. (2024). Journal of Experimental Botany, erae451.
SlMIPS2, a myo-inositol phosphate synthase, regulates phosphate homeostasis by influencing SlPHL1 and SlSPX2 levels in tomato seedlings.
Roychowdhury, A., Kaushik, D., Bhosale, J. B., Srivastava, R. & Kumar, R. (2024). Journal of Experimental Botany, erae451.
Phosphorus (P) is a quintessential macronutrient utilized by plants to support various metabolic processes during growth and development. Recent studies have revealed the pivotal role of inositol hexa-kis/pyrophosphate (InsP6-8), the derivatives of Myo-inositol (MI), in facilitating the interaction between SYG1/PHO81/XPR1 (SPX) and Phosphate starvation response (PHR) proteins. Myo-inositol phosphate synthase (MIPS) catalyzes the first committed step in MI biosynthesis. Although the role of MIPS genes in mediating stress responses in plants is well elucidated, its role in phosphate (Pi) deficiency remains largely unexplored. This study demonstrates that out of the five MIPS genes encoded by the tomato genome, only SlMIPS2 is sharply induced at an early stage of Pi starvation in tomato seedlings. Silencing of SlMIPS2 led to improved seedling growth with enhanced total soluble Pi and total P levels in the silenced plants under high Pi availability. SlMIPS2 silencing also caused a significant reduction in MI and InsP6 content in the tomato seedlings. These seedlings with depleted InsP6 levels accumulated lower levels of SlSPX2 protein. In contrast, stabilized SlPHL1 levels were noticed in these plants, directly implicating this transcription factor in activating phosphate starvation inducible (PSI) genes in the SlMIPS2-silenced seedlings, even under high Pi conditions. The results assign a novel role to SlMIPS2 in regulating cellular InsP6 levels and SPX-PHR interactions to control Pi homeostasis in tomato seedlings.
Inositol in Disease and Development: Roles of Catabolism via myo-Inositol Oxygenase in Drosophila melanogaster.
Contreras, A., Jones, M. K., Eldon, E. D. & Klig, L. S. (2023). International Journal of Molecular Sciences, 24(4), 4185.
Inositol in Disease and Development: Roles of Catabolism via myo-Inositol Oxygenase in Drosophila melanogaster.
Contreras, A., Jones, M. K., Eldon, E. D. & Klig, L. S. (2023). International Journal of Molecular Sciences, 24(4), 4185.
Inositol depletion has been associated with diabetes and related complications. Increased inositol catabolism, via myo-inositol oxygenase (MIOX), has been implicated in decreased renal function. This study demonstrates that the fruit fly Drosophila melanogaster catabolizes myo-inositol via MIOX. The levels of mRNA encoding MIOX and MIOX specific activity are increased when fruit flies are grown on a diet with inositol as the sole sugar. Inositol as the sole dietary sugar can support D. melanogaster survival, indicating that there is sufficient catabolism for basic energy requirements, allowing for adaptation to various environments. The elimination of MIOX activity, via a piggyBac WH-element inserted into the MIOX gene, results in developmental defects including pupal lethality and pharate flies without proboscises. In contrast, RNAi strains with reduced levels of mRNA encoding MIOX and reduced MIOX specific activity develop to become phenotypically wild-type-appearing adult flies. myo-Inositol levels in larval tissues are highest in the strain with this most extreme loss of myo-inositol catabolism. Larval tissues from the RNAi strains have inositol levels higher than wild-type larval tissues but lower levels than the piggyBac WH-element insertion strain. myo-Inositol supplementation of the diet further increases the myo-inositol levels in the larval tissues of all the strains, without any noticeable effects on development. Obesity and blood (hemolymph) glucose, two hallmarks of diabetes, were reduced in the RNAi strains and further reduced in the piggyBac WH-element insertion strain. Collectively, these data suggest that moderately increased myo-inositol levels do not cause developmental defects and directly correspond to reduced larval obesity and blood (hemolymph) glucose.
Myo-Inositol Supplementation in Suckling Rats Protects against Adverse Programming Outcomes on Hypothalamic Structure Caused by Mild Gestational Calorie Restriction, Partially Comparable to Leptin Effects.
Castillo, P., Palou, M., Yau-Qiu, Z. X., Rodríguez, A. M., Palou, A. & Picó, C. (2021). Nutrients, 13(9), 3257.
Myo-Inositol Supplementation in Suckling Rats Protects against Adverse Programming Outcomes on Hypothalamic Structure Caused by Mild Gestational Calorie Restriction, Partially Comparable to Leptin Effects.
Castillo, P., Palou, M., Yau-Qiu, Z. X., Rodríguez, A. M., Palou, A. & Picó, C. (2021). Nutrients, 13(9), 3257.
We studied whether myo-inositol supplementation throughout lactation, alone and combined with leptin, may reverse detrimental effects on hypothalamic structure and function caused by gestational calorie gestation (CR) in rats. Candidate early transcript-based biomarkers of metabolic health in peripheral blood mononuclear cells (PBMC) were also studied. Offspring of dams exposed to 25% gestational CR and supplemented during lactation with physiological doses of leptin (CR-L), myo-inositol (CR-M), the combination (CR-LM), or the vehicle (CR-V) as well as control rats (CON-V) were followed and sacrificed at postnatal day 25. Myo-inositol and the combination increased the number of neurons in arcuate nucleus (ARC) (only in females) and paraventricular nucleus, and myo-inositol (alone) restored the number of αMSH+ neurons in ARC. Hypothalamic mRNA levels of Lepr in CR-M and Insr in CR-M and CR-LM males were higher than in CR-V and CON-V, respectively. In PBMC, increased expression levels of Lrp11 and Gls in CR-V were partially normalized in all supplemented groups (but only in males for Gls). Therefore, myo-inositol supplementation throughout lactation, alone and combined with leptin, reverts programmed alterations by fetal undernutrition on hypothalamic structure and gene expression of potential early biomarkers of metabolic health in PBMC, which might be attributed, in part, to increased leptin sensitivity.
Deletion of Voltage-Dependent Anion Channel 1 knocks mitochondria down triggering metabolic rewiring in yeast.
Magri, A., Di Rosa, M. C., Orlandi, I., Guarino, F., Reina, S., Guarnaccia, M., Morello, G., Spampinato, A., Cavallaro, S., Messina, A., Vai, M. & De Pinto, V. (2020). Cellular and Molecular Life Sciences, 77(16), 3195-3213.
Deletion of Voltage-Dependent Anion Channel 1 knocks mitochondria down triggering metabolic rewiring in yeast.
Magri, A., Di Rosa, M. C., Orlandi, I., Guarino, F., Reina, S., Guarnaccia, M., Morello, G., Spampinato, A., Cavallaro, S., Messina, A., Vai, M. & De Pinto, V. (2020). Cellular and Molecular Life Sciences, 77(16), 3195-3213.
The Voltage-Dependent Anion-selective Channel (VDAC) is the pore-forming protein of mitochondrial outer membrane, allowing metabolites and ions exchanges. In Saccharomyces cerevisiae, inactivation of POR1, encoding VDAC1, produces defective growth in the presence of non-fermentable carbon source. Here, we characterized the whole-genome expression pattern of a VDAC1-null strain (Δpor1) by microarray analysis, discovering that the expression of mitochondrial genes was completely abolished, as consequence of the dramatic reduction of mtDNA. To overcome organelle dysfunction, Δpor1 cells do not activate the rescue signaling retrograde response, as ρ0 cells, and rather carry out complete metabolic rewiring. The TCA cycle works in a “branched” fashion, shunting intermediates towards mitochondrial pyruvate generation via malic enzyme, and the glycolysis-derived pyruvate is pushed towards cytosolic utilization by PDH bypass rather than the canonical mitochondrial uptake. Overall, Δpor1 cells enhance phospholipid biosynthesis, accumulate lipid droplets, increase vacuoles and cell size, overproduce and excrete inositol. Such unexpected re-arrangement of whole metabolism suggests a regulatory role of VDAC1 in cell bioenergetics.
Impaired GCR1 transcription resulted in defective inositol levels, vacuolar structure and autophagy in Saccharomyces cerevisiae.
Ravi, C., Gowsalya, R. & Nachiappan, V. (2019). Current Genetics, 65(4), 995-1014.
Impaired GCR1 transcription resulted in defective inositol levels, vacuolar structure and autophagy in Saccharomyces cerevisiae.
Ravi, C., Gowsalya, R. & Nachiappan, V. (2019). Current Genetics, 65(4), 995-1014.
In yeast, the GCR1 transcription factor is involved in the regulation of glycolysis and its deletion exhibited growth defect, reduced inositol and phosphatidylinositol (PI) levels compared to WT cells. We observed a down regulation of the INO1 and PIS1 expression in gcr1∆ cells under both I- and I+ conditions and the over expression of GCR1 in gcr1∆ cells restored the growth, retrieved the expression of INO1, and PIS1 comparable to WT cells. In the gel shift assay, the Gcr1p binds to its consensus sequence CTTCC in PIS1 promoter and regulates its expression but not in INO1 transcription. The WT cells, under I- significantly reduced the expression of GCR1 and PIS1, but increased the expression of KCS1 and de-repressed INO1. The Kcs1p expression was reduced in gcr1∆ cells; this reduced INO1 expression resulting in abnormal vacuolar structure and reduced autophagy in Saccharomyces cerevisiae.
Ivanov Kavkova, E., Blöchl, C. & Tenhaken, R. (2018). Plant Biology, In Press.
Ivanov Kavkova, E., Blöchl, C. & Tenhaken, R. (2018). Plant Biology, In Press.
Ascorbic acid (AsA) biosynthesis in plants predominantly occurs via a pathway with D‐mannose and L‐galactose as intermediates. One alternative pathway for AsA synthesis, which is similar to the biosynthesis route in mammals, is controversially discussed for plants. Here, myo‐inositol is cleaved to glucuronic acid and then converted via L‐gulonate to AsA. In contrast to animals, plants have an effective recycling pathway for glucuronic acid being a competitor for the metabolic rate. Recycling involves a phosphorylation at C1 by the enzyme glucuronokinase. Two previously described T‐DNA insertion lines in the gene coding for glucuronokinase1 show wildtype like expression levels of the mRNA in our experiments and do not accumulate glucuronic acid in labelling experiments disproving that these lines are true knockouts. As suitable T‐DNA insertion lines were not available, we generated frameshift mutations in the major expressed isoform glucuronokinase1 (At3g01640) to potentially redirect metabolites to AsA. However, radiotracer experiments with 3Hmyo‐inositol revealed that the mutants in glucuronokinase1 accumulate only glucuronic acid and incorporate less metabolite into cell wall polymers. AsA was not labeled, suggesting that Arabidopsis cannot efficiently use glucuronic acid for AsA biosynthesis. All four mutants in glucuronokinase as well as wildtype have the same level of AsA in leaves.
Brüsseler, C., Radek, A., Tenhaef, N., Krumbach, K., Noack, S. & Marienhagen, J. (2017). Bioresource Technology, 249, 953-961.
Brüsseler, C., Radek, A., Tenhaef, N., Krumbach, K., Noack, S. & Marienhagen, J. (2017). Bioresource Technology, 249, 953-961.
Corynebacterium glutamicum has been engineered to utilize D-xylose as sole carbon and energy source. Recently, a C. glutamicum strain has been optimized for growth on defined medium containing D-xylose by laboratory evolution, but the mutation(s) attributing to the improved-growth phenotype could not be reliably identified. This study shows that loss of the transcriptional repressor IolR is responsible for the increased growth performance on defined D-xylose medium in one of the isolated mutants. Underlying reason is derepression of the gene for the glucose/myo-inositol permease IolT1 in the absence of IolR, which could be shown to also contribute to D-xylose uptake in C. glutamicum. IolR-regulation of iolT1 could be successfully repealed by rational engineering of an IolR-binding site in the iolT1-promoter. This minimally engineered C. glutamicum strain bearing only two nucleotide substitutions mimics the IolR loss-of-function phenotype and allows for a high growth rate on D-xylose-containing media (µmax).= 0.24 ± 0.01 h-1).
Takeno, S., Hori, K., Ohtani, S., Mimura, A., Mitsuhashi, S. & Ikeda, M. (2016). Metabolic engineering, 37, 1-10.
Takeno, S., Hori, K., Ohtani, S., Mimura, A., Mitsuhashi, S. & Ikeda, M. (2016). Metabolic engineering, 37, 1-10.
We have recently developed a Corynebacterium glutamicum strain that generates NADPH via the glycolytic pathway by replacing endogenous NAD-dependent glyceraldehyde 3-phosphate dehydrogenase (GapA) with a nonphosphorylating NADP-dependent glyceraldehyde 3-phosphate dehydrogenase (GapN) from Streptococcus mutans. Strain RE2, a suppressor mutant spontaneously isolated for its improved growth on glucose from the engineered strain, was proven to be a high-potential host fof L-lysine production (Takeno et al., 2010). In this study, the suppressor mutation was identified to be a point mutation in rho encoding the transcription termination factor Rho. Strain RE2 still showed retarded growth despite the mutation rho696. Our strategy for reconciling improved growth with a high level of L-lysine production was to use GapA together with GapN only in the early growth phase, and subsequently shift this combination-type glycolysis to one that depends only on GapN in the rest of the growth phase. To achieve this, we expressed gapA under the myo-inositol-inducible promoter of iolT1 encoding a myo-inositol transporter in strain RE2. The resulting strain RE2Aiol was engineered into an L-lysine producer by introduction of a plasmid carrying the desensitized lysC, followed by examination for culture conditions with myo-inositol supplementation. We found that as a higher concentration of myo-inositol was added to the seed culture, the following fermentation period became shorter while maintaining a high level of L-lysine production. This finally reached a fermentation period comparable to that of the control GapA strain, and yielded a 1.5-fold higher production rate compared with strain RE2. The transcript level of gapA, as well as the GapA activity, in the early growth phase increased in proportion to the myo-inositol concentration and then fell to low levels in the subsequent growth phase, indicating that improved growth was a result of increased GapA activity, especially in the early growth phase. Moreover, blockade of the pentose phosphate pathway through a defect in glucose 6-phosphate dehydrogenase did not significantly affect L-lysine production in the engineered GapN strains, while a drastic decrease in L-lysine production was observed for the control GapA strain. Determination of the intracellular NADPH/NADP+ ratios revealed that the ratios in the engineered strains were significantly higher than the ratio of the control GapA strain irrespective of the pentose phosphate pathway. These results demonstrate that our strain engineering strategy allows efficient L-lysine production independent of the oxidative pentose phosphate pathway.
Koguchi, T., Tanikawa, C., Mori, J., Kojima, Y. & Matsuda, K. (2016). International Journal of Oncology, 48(6), 2415-2424.
Koguchi, T., Tanikawa, C., Mori, J., Kojima, Y. & Matsuda, K. (2016). International Journal of Oncology, 48(6), 2415-2424.
In response to various cellular stresses, p53 exerts its tumor suppressive effects such as apoptosis, cell cycle arrest, and senescence through the induction of its target genes. Recently, p53 was shown to control cellular homeostasis by regulating energy metabolism, glycolysis, antioxidant effect, and autophagy. However, its function in inositol synthesis was not reported. Through a microarray screening, we found that five genes related with myo-inositol metabolism were induced by p53. DNA damage enhanced intracellular myo-inositol content in HCT116 p53+/+ cells, but not in HCT116 p53-/- cells. We also indicated that inositol 3-phosphate synthase (ISYNA1) which encodes an enzyme essential for myo-inositol biosynthesis as a direct target of p53. Activated p53 regulated ISYNA1 expression through p53 response element in the seventh exon. Ectopic ISYNA1 expression increased myo-inositol levels in the cells and suppressed tumor cell growth. Knockdown of ISYNA1 caused resistance to adriamycin treatment, demonstrating the role of ISYNA1 in p53-mediated growth suppression. Furthermore, ISYNA1 expression was significantly associated with p53 mutation in bladder, breast cancer, head and neck squamous cell carcinoma, lung squamous cell carcinoma, and pancreatic adenocarcinoma. Our findings revealed a novel role of p53 in myo-inositol biosynthesis which could be a potential therapeutic target.
Safety Information
Download Safety Data Sheet
Select a specific region and the Safety Data Sheet will open in a new window. Please make sure your browser doesn’t block pop-up windows

Receive Megazyme’s latest articles, news, product introductions and promotions
Customers also viewed
 Assay Kit (K-HDBA)")
60 assays (manual) / 600 assays (microplate) / 740 assays (auto-analyser)
")
70 assays (manual) / 700 assays (microplate) / 600 assays (auto-analyser)
 (K-ACET)")
 (K-CERA)")
")
Total Dietary Fiber Assay Kit
700004347 / K-TDFR-200A200 assays
Total Dietary Fiber Assay Kit
700004346 / K-TDFR-100A100 assays
")



