 Assay Kit (K-HDBA)")
 Assay Kit (K-HDBA)")
D-3-Hydroxybutyric Acid (β-Hydroxybutyrate) Assay Kit
60 assays (manual) / 600 assays (microplate) / 740 assays (auto-analyser)
Long term stability: See individual component labels
Acid
Advantages
- Very competitive price (cost per test)
- All reagents stable for > 2 years after preparation
- Very rapid reaction (~ 3 min)
- No wasted diaphorase solution (stable suspension supplied)
- Mega-Calc™ software tool is available from our website for hassle-free raw data processing
- Standard included
- Suitable for manual, microplate and auto-analyser formats
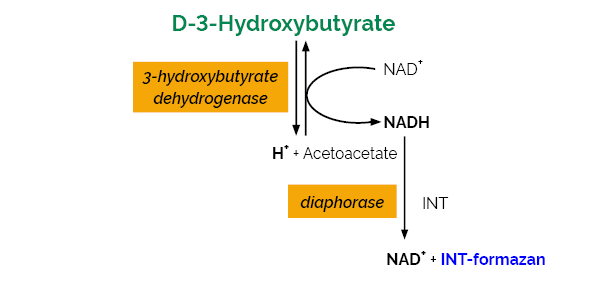
The D-3-Hydroxybutyric Acid (β-Hydroxybutyrate) Assay Kit is suitable for the specific measurement and analysis of D-hydroxybutyric acid in eggs and egg products and other foods and beverages.
Note for Content: The number of manual tests per kit can be doubled if all volumes are halved. This can be readily accommodated using the MegaQuantTM Wave Spectrophotometer (D-MQWAVE).
Explore more organic acid assay kit products.
Publications
Deciphering heterogeneous enzymatic surface reactions on xylan using surface plasmon resonance spectroscopy.
Schaubeder, J. B., Fürk, P., Amering, R., Gsöls, L., Ravn, J., Nypelö, T. & Spirk, S. (2024). Carbohydrate Polymers, 337, 122137.
Deciphering heterogeneous enzymatic surface reactions on xylan using surface plasmon resonance spectroscopy.
Schaubeder, J. B., Fürk, P., Amering, R., Gsöls, L., Ravn, J., Nypelö, T. & Spirk, S. (2024). Carbohydrate Polymers, 337, 122137.
Xylans' unique properties make it attractive for a variety of industries, including paper, food, and biochemical production. While for some applications the preservation of its natural structure is crucial, for others the degradation into monosaccharides is essential. For the complete breakdown, the use of several enzymes is required, due to its structural complexity. In fact, the specificity of enzymatically-catalyzed reactions is guided by the surface, limiting or regulating accessibility and serving structurally encoded input guiding the actions of the enzymes. Here, we investigate enzymes at surfaces rich in xylan using surface plasmon resonance spectroscopy. The influence of diffusion and changes in substrate morphology is studied via enzyme surface kinetics simulations, yielding reaction rates and constants. We propose kinetic models, which can be applied to the degradation of multilayer biopolymer films. The most advanced model was verified by its successful application to the degradation of a thin film of polyhydroxybutyrate treated with a polyhydroxybutyrate-depolymerase. The herein derived models can be employed to quantify the degradation kinetics of various enzymes on biopolymers in heterogeneous environments, often prevalent in industrial processes. The identification of key factors influencing reaction rates such as inhibition will contribute to the quantification of intricate dynamics in complex systems.
Fermentation Quality and Chemical Composition of Industrial Hemp (Cannabis sativa L.) Silage Inoculated with Bacterial Starter Cultures-A Pilot Study.
Wróbel, B., Hryniewicz, M., Kulkova, I., Mazur, K., Jakubowska, Z., Borek, K., Dobrzyński, J., Konieczna, A., Miecznikowski, A., Piasecka-Jóźwiak, K. & Fabiszewska, A. (2023). Agronomy, 13(5), 1371.
Fermentation Quality and Chemical Composition of Industrial Hemp (Cannabis sativa L.) Silage Inoculated with Bacterial Starter Cultures-A Pilot Study.
Wróbel, B., Hryniewicz, M., Kulkova, I., Mazur, K., Jakubowska, Z., Borek, K., Dobrzyński, J., Konieczna, A., Miecznikowski, A., Piasecka-Jóźwiak, K. & Fabiszewska, A. (2023). Agronomy, 13(5), 1371.
Industrial hemp (Cannabis sativa L.) is a plant species cultivated as a raw material for fiber extraction. Alternatively, hemp biomass can be used for feeding or energy purposes. This study was conducted to investigate the effect of inoculation with a lactic acid bacteria starter culture on the fermentation and chemical compositions of hemp silages. Hemp shoots (HS) and hemp flowers (HF) were ensiled in mini laboratory silos without or with the inoculation of the commercial starter culture Lactosil Biogaz (Lentilactobacillus buchnerii KKP 907 p; L. buchneri A KKP 2047 p; Pediococcus acidilactici KKP 2065 p). After 7 and 42 days of ensiling, the fermentation quality and chemical compositions of the silages were assessed. The use of Lactosil Biogas for hemp resulted in a decrease in pH, increase in lactic acid (LA), and reduction in fungal abundance in the HS silage. In the case of the HF silage, the bacterial inoculation was less effective; however, an increase in LA and a decrease in butyric acid (BA) were observed. As a result of the ensilage process, decreases in crude fiber and hemicellulose were observed in the HS and HF silages. Thus, hemp ensiling with biological additives is an effective pre-treatment of hemp plants for subsequent biofuel production that can preserve the biomass and provide the year-round availability of feedstock.
Systems biology engineering of the pantothenate pathway to enhance 3HB productivity in Escherichia coli.
Younes, S., Awad, D., Kassab, E., Haack, M., Schuler, C., Mehlmer, N. & Brueck, T. (2021). Biotechnology and Bioprocess Engineering, 26(4), 621-629.
Systems biology engineering of the pantothenate pathway to enhance 3HB productivity in Escherichia coli.
Younes, S., Awad, D., Kassab, E., Haack, M., Schuler, C., Mehlmer, N. & Brueck, T. (2021). Biotechnology and Bioprocess Engineering, 26(4), 621-629.
The monomer, 3-hydroxybutyrate (3HB), plays an interesting role as a precursor for antibiotics, vitamins, and bioplastics such as polyhydroxybutyrates (PHB). The biotechnological production of both compounds in Escherichia coli has seen increased interest in the last decade. The central metabolite in the 3HB production pathway is acetyl-CoA, the derivative of coenzyme A (CoA). Enriching the intracellular pool of these cofactors could improve 3HB titers. In our study, we opted to increase CoA titers by upregulating pantothenate kinase (PanK), the rate limiting step in CoA biosynthetic pathway. To this end, 4 PanKs genes of different taxonomic origins (mammalian, fungal, and bacterial) were individually expressed and evaluated in 3HB producing E. coli cells. In shake flask studies, strains expressing Aspergillus nidulans PankII and Mus musculus PanK1β achieved the highest 3HB titers. In a bioreactor fermentation, the strain harboring murine PanK1β resulted in 7.6 g/L compared to 5.4 g/L of 3HB in the control strain. Although structurally different from the bacterial PankI, our study showed that eukaryotic Panks can supplement the kinase activity in prokaryotes. Expressing the exogenous PanKs offer several advantages over the host’s enzyme; PanKII is only inhibited by acetyl-CoA, for which the 3HB-production system would provide a constant metabolic sink. Additionally, PanK1β is weakly regulated by acetyl-CoA, and its activity is stimulated by free CoA. Overexpressing eukaryotic PanKs constitutes a suitable strategy for improving 3HB titers in E. coli.
NIK links inflammation to hepatic steatosis by suppressing PPARα in alcoholic liver disease.
Li, Y., Chen, M., Zhou, Y., Tang, C., Zhang, W., Zhong, Y., Chen, Y., Zhou, H. & Sheng, L. (2020). Theranostics, 10(8), 3579.
NIK links inflammation to hepatic steatosis by suppressing PPARα in alcoholic liver disease.
Li, Y., Chen, M., Zhou, Y., Tang, C., Zhang, W., Zhong, Y., Chen, Y., Zhou, H. & Sheng, L. (2020). Theranostics, 10(8), 3579.
Background: Inflammation and steatosis are the main pathological features of alcoholic liver disease (ALD), in which, inflammation is one of the critical drivers for the initiation and development of alcoholic steatosis. NIK, an inflammatory pathway component activated by inflammatory cytokines, was suspected to link inflammation to hepatic steatosis during ALD. However, the underlying pathogenesis is not well-elucidated. Methods: Alcoholic steatosis was induced in mice by chronic-plus-binge ethanol feeding. Both the loss- and gain-of-function experiments by the hepatocyte-specific deletion, pharmacological inhibition and adenoviral transfection of NIK were utilized to elucidate the role of NIK in alcoholic steatosis. Rate of fatty acid oxidation was assessed in vivo and in vitro. PPARα agonists or antagonists of MEK1/2 and ERK1/2 were used to identify the NIK-induced regulation of PPARα, MEK1/2, and ERK1/2. The potential interactions between NIK, MEK1/2, ERK1/2 and PPARα and the phosphorylation of PPARα were clarified by immunoprecipitation, immunoblotting and far-western blotting analysis. Results: Hepatocyte-specific deletion of NIK protected mice from alcoholic steatosis by sustaining hepatic fatty acid oxidation. Moreover, overexpression of NIK contributed to hepatic lipid accumulation with disrupted fatty acid oxidation. The pathological effect of NIK in ALD may be attributed to the suppression of PPARα, the main controller of fatty acid oxidation in the liver, because PPARα agonists reversed NIK-mediated hepatic steatosis and malfunction of fatty acid oxidation. Mechanistically, NIK recruited MEK1/2 and ERK1/2 to form a complex that catalyzed the inhibitory phosphorylation of PPARα. Importantly, pharmacological intervention against NIK significantly attenuated alcoholic steatosis in ethanol-fed mice. Conclusions: NIK targeting PPARα via MEK1/2 and ERK1/2 disrupts hepatic fatty acid oxidation and exhibits high value in ALD therapy.
Thiel, K., Vuorio, E., Aro, E. M. & Kallio, P. T. (2017). Microbial Cell Factories, 16(1), 21.
Thiel, K., Vuorio, E., Aro, E. M. & Kallio, P. T. (2017). Microbial Cell Factories, 16(1), 21.
Background: Acetate is a common microbial fermentative end-product, which can potentially be used as a supplementary carbon source to enhance the output of biotechnological production systems. This study focuses on the acetate metabolism of the photosynthetic cyanobacterium Synechocystis sp. PCC 6803 which is unable to grow on acetate as a sole carbon source but still can assimilate it via acetyl-CoA—derived metabolic intermediates. In order to gain insight into the acetate uptake, associated limitations and metabolic effects, a heterologous acetate transporter ActP from Escherichia coli was introduced into Synechocystis to facilitate the transport of supplemented acetate from the medium into the cell. Results: The results show that enhanced acetate intake can efficiently promote the growth of the cyanobacterial host. The effect is apparent specifically under low-light conditions when the photosynthetic activity is low, and expected to result from increased availability of acetyl-CoA precursors, accompanied by changes induced in cellular glycogen metabolism which may include allocation of resources towards enhanced growth instead of glycogen accumulation. Despite the stimulated growth of the mutant, acetate is shown to suppress the activity of the photosynthetic apparatus, further emphasizing the contribution of glycolytic metabolism in the acetate-induced effect. Conclusions: The use of acetate by the cyanobacterium Synechocystis sp. PCC 6803 is at least partially restricted by the import into the cell. This can be improved by the introduction of a heterologous acetate transporter into the system, thereby providing a potential advantage by expanding the scope of acetate utilization for various biosynthetic processes.
Wagner, D. N., Green, D. J., Pavlik, M., Cooper, J. & Williams, T. D. (2014). Conservation Physiology, 2(1), cou017.
Wagner, D. N., Green, D. J., Pavlik, M., Cooper, J. & Williams, T. D. (2014). Conservation Physiology, 2(1), cou017.
Riparian habitat makes up a small fraction of the landscape but provides important stopover habitat for migratory birds. Hydroelectric dam operations cause fluctuations in water levels that can change the amount or quality of riparian habitat, which in turn might affect potential fattening rates of migrant birds. Here we used plasma metabolite analysis to estimate variation in fattening rate in relationship to variable water levels associated with reservoir management in four species of neotropical migratory songbirds using riparian habitat at a dam-impacted stopover site in Revelstoke, British Columbia, Canada. Residual plasma triglyceride, our measure of estimated fattening rate, varied systematically with time of day and Julian date and varied consistently among species, but did not vary with age or sex. Controlling for potentially confounding variables, we found no inter-annual variation in estimated fattening rate, even though there were marked differences in water levels among years. Likewise, there was no relationship between daily variation in water levels and estimated fattening rate. Data on feather isotopes (δD), indicative of migratory origin, did not add explanatory power to our models. There was inter-annual variation in plasma glycerol and β-hydroxybutyrate levels and significant, though weak, relationships between these metabolites and water level (higher metabolite levels when drier) that might indicate effects on ‘body condition’ independent of fattening rate. Our study suggests that, at present, although hydroelectric dam operations influence water levels in the Arrows Lake Reservoir and adjacent riparian habitats, this does not significantly impact fattening rates of migratory passerines using these habitats.
Safety Information
Download Safety Data Sheet
Select a specific region and the Safety Data Sheet will open in a new window. Please make sure your browser doesn’t block pop-up windows

Receive Megazyme’s latest articles, news, product introductions and promotions
Customers also viewed
 (K-MRH2O2)")
1000 assays (microplate) / 100 assays (manual) / 400 assays (auto-analyser)
")
200 assays (manual) / 2000 assays (microplate) / 1960 assays (auto-analyser)
")
")
")
60 assays (manual) / 600 assays (microplate) / 700 assays (auto-analyser)
 (K-LATELQ)")



